
Travaux de recherche
sur le fonctionnement hydraulique des arbres
UMR 547
PIAF, INRA-UBP. Equipe HYDRO
Contact : Hervé Cochard (cochard@clermont.inra.fr)
Pour des nouvelles plus fraiches de mes recherches, suivez
moi maintenant sur Twitter @HCochard
Décembre 2010 : Lancement de l’ANR ‘PitBulles’
 Projet
ANR Blanc Interdisciplinaire PitBulles (2011-2014) “Déterminismes
biophysiques et moléculaires de la vulnérabilité à l'embolie chez les arbres »
Projet
ANR Blanc Interdisciplinaire PitBulles (2011-2014) “Déterminismes
biophysiques et moléculaires de la vulnérabilité à l'embolie chez les arbres »
Coordination : Hervé Cochard (UMR Physique et physiologie
Intégratives des Arbres Forestiers ou
fruitiers (PIAF), INRA Université Blaise Pascal, Crouel)
Les arbres irriguent leurs organes par un système de
transfert d’eau qui opère sous des pressions hydrostatiques très négatives,
particulièrement lorsqu’ils sont exposés à des contraintes hydriques. Dans ces
conditions métastables, les arbres vivent sous le risque d’une vaporisation
soudaine de leur sève, l’embolie. Les travaux effectués au cours des deux
dernières décennies sur le fonctionnement hydraulique des arbres ont conduit à
la conclusion importante que la résistance à l’embolie est un caractère
adaptatif majeur de la tolérance à la sécheresse. Cependant, les travaux sur la
diversité génétique et les implications écologiques de ce trait demeurent
encore très limités, pour deux raisons. Tout d’abord, certains travaux étaient
inenvisageables car aucune technique ne permettait l’étude de ce caractère sur
un grand nombre d’individus. Le développement de la technique du « Cavitron »
par l’équipe porteuse de ce projet a levé récemment ce verrou méthodologique.
En revanche, les bases génétiques de la résistance à l’embolie restent
inconnues à ce jour, ce qui freine considérablement les recherches dans les
domaines de l’écologie moléculaire ou de la génétique des populations.
L’ambition de ce projet est de lever ce second verrou.
La démarche que nous avons mise en place consiste à définir les structures
anatomiques déterminantes pour la formation de l’embolie dans l’objectif
d’identifier les gènes codants pour ces structures. Dans l’état actuel des
connaissances, l’embolie se propage dans le tissu vasculaire des arbres
lorsqu’une bulle d’air pénètre à travers les parois des conduits, au niveau des
ponctuations, structure anatomique permettant le passage de la sève d’un
conduit à l’autre. L’hypothèse principale qui sous-tend ce projet est que le
mécanisme d’embolie est déterminé par la structure et les propriétés
physico-chimiques de la paroi primaire des ponctuations. Une partie centrale de
ce projet consistera à analyser ces propriétés pour des espèces, des génotypes
et des phénotypes contrastés vis-à-vis de leur vulnérabilité à l’embolie. Une
attention particulière sera portée sur la composition pectique de ces parois
car des résultats préliminaires attestent de leur rôle majeur dans ce
mécanisme. Ces travaux sur la structure fine des ponctuations formeront la base
d’une analyse micro-fluidique de leur fonctionnement. Il s’agira ici de
proposer un modèle physique explicatif du mécanisme d’embolie. Ce modèle sera
validé par des expérimentations sur des parois biomimétiques artificielles. A
l’issue de ces investigations, nous seront en mesure de proposer une liste
ciblée de gènes codants spécifiquement pour les structures clés des
ponctuations et putativement impliqués dans la
résistance à l'embolie. Le dernier volet de ce projet consistera à valider
l’implication de ces gènes grâce aux outils de la biologie moléculaires.
Le projet repose sur l’interdisciplinarité et complémentarité de trois
partenaires, leaders chacun dans des thématiques clés pour cette étude :
hydraulique des arbres (équipe Hydro, UMR PIAF), parois primaires des végétaux
(Unité BIA, INRA-Nantes), mécanique des fluides diphasiques (IMFT, CNRS
Toulouse). Cette démarche interdisciplinaire proposée ici pour la première fois
devrait permettre de lever des verrous scientifiques majeurs et ainsi ouvrir
les portes vers des recherches innovantes sur les ressources génétiques
forestières.
Aout
2010 : Mise en évidence du rôle du Calcium dans la résistance des arbres à
la cavitation.
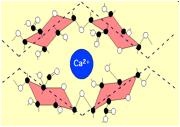 La
vulnérabilité à la cavitation du xylème est un paramètre clé dans la tolérance
à la sécheresse des arbres, mais on sait peu sur les mécanismes moléculaires
impliqués dans ce processus. La cavitation est supposée se produire lorsqu’une
bulle d'air pénètre à travers la paroi primaire d’une ponctuation. Elle serait
donc influencée par la structure et
la texture de cette paroi. Dans ce travail, nous avons testé le rôle du calcium
présent dans cette paroi sur la vulnérabilité à la cavitation du Hêtre. Des tiges perfusées avec des solutions
d'acide oxalique, EGTA, ou du phosphate de sodium (NaPO4) sont plus rendues
beaucoup vulnérables à la cavitation.
L’augmentation de la vulnérabilité à la cavitation induite par le NaPO4 est
liée à l'élimination du calcium de cette de la paroi. En revanche, la conductance hydraulique du xylème n'a
pas été affectée par les traitements chimiques, ce qui démontre que les
mécanismes de contrôle de la vulnérabilité à la cavitation et de la
résistance hydraulique sont découplés. L’effet du NaPO4 a ensuite été testé sur
13 espèces d'arbres possédant des
vulnérabilités très contrastées à la cavitation. Le calcium se révèle être un
déterminant majeur de la différence de vulnérabilité à la cavitation entre ces
espèces. Ce rôle a été démontré tant les angiospermes chez es espèces de
conifères, soutenant ainsi l'hypothèse
d'un mécanisme commun de la sécheresse induite par cavitation. Le projet ‘PitBulles’
financé par l’Agence Nationale de la Recherche nous permettra d’explorer plus
encore les bases génétiques et biophysique de la cavitation des Arbres.
La
vulnérabilité à la cavitation du xylème est un paramètre clé dans la tolérance
à la sécheresse des arbres, mais on sait peu sur les mécanismes moléculaires
impliqués dans ce processus. La cavitation est supposée se produire lorsqu’une
bulle d'air pénètre à travers la paroi primaire d’une ponctuation. Elle serait
donc influencée par la structure et
la texture de cette paroi. Dans ce travail, nous avons testé le rôle du calcium
présent dans cette paroi sur la vulnérabilité à la cavitation du Hêtre. Des tiges perfusées avec des solutions
d'acide oxalique, EGTA, ou du phosphate de sodium (NaPO4) sont plus rendues
beaucoup vulnérables à la cavitation.
L’augmentation de la vulnérabilité à la cavitation induite par le NaPO4 est
liée à l'élimination du calcium de cette de la paroi. En revanche, la conductance hydraulique du xylème n'a
pas été affectée par les traitements chimiques, ce qui démontre que les
mécanismes de contrôle de la vulnérabilité à la cavitation et de la
résistance hydraulique sont découplés. L’effet du NaPO4 a ensuite été testé sur
13 espèces d'arbres possédant des
vulnérabilités très contrastées à la cavitation. Le calcium se révèle être un
déterminant majeur de la différence de vulnérabilité à la cavitation entre ces
espèces. Ce rôle a été démontré tant les angiospermes chez es espèces de
conifères, soutenant ainsi l'hypothèse
d'un mécanisme commun de la sécheresse induite par cavitation. Le projet ‘PitBulles’
financé par l’Agence Nationale de la Recherche nous permettra d’explorer plus
encore les bases génétiques et biophysique de la cavitation des Arbres.
Publication : Herbette S, Cochard
H (2010) Calcium
is a major determinant of xylem vulnerability to cavitation. Plant
Physiology 153: 1932–1939. pdf
Avril 2010 : Une faille dans les techniques de
mesure de la cavitation.
 Dans les plantes,
la sève circule sous tension, c'est-à-dire sous des pressions négatives !
Cette état de l’eau est physiquement instable et peut évoluer vers une phase
gazeuse, plus stable. C’est le phénomène de cavitation. Il provoque une embolie
vasculaire qui bloque la circulation de la sève. C’est une perturbation majeure
car elle peut conduire à la mort des plantes (voir la brève de Janvier 2009
ci-dessous). De nombreuses techniques ont été proposées pour étudier la
cavitation et l’embolie des plantes et notre laboratoire a été particulièrement
pionnier dans ce domaine. Les techniques ‘anciennes’ sont fiables, mais peu
rapides. Elles ont néanmoins permis de faire considérablement progresser notre
compréhension du fonctionnement hydrique des plantes (dans le domaine c’est
probablement l’une des avancées majeures des 2 dernières décennies, avec la
découverte des aquaporines). Les
résultats acquis avec ces méthodes suggèrent que la cavitation (estivale) est
un phénomène exceptionnel qui se produit lorsque les plantes atteignent leur
limite de fonctionnement, par exemple lors d’une sécheresse d’une grande
ampleur (1976, 2003). On pense donc que c’est un processus impliqué dans
la survie ou la résilience des espèces à ces sécheresses. Les techniques
‘modernes’ (telle la centrifugation ou l’injection d’air) sont beaucoup plus
rapides. Certaines, comme le ‘Cavitron’ que nous avons développé au
laboratoire, permettent même de faire du phénotypage
à moyen débit de la cavitation. Ces techniques sont maintenant utilisées en
routine dans de nombreux laboratoires dans le monde. Pour certaines espèces,
les résultats obtenus avec ces techniques changent radicalement notre
perception du phénomène de cavitation. En effet, certaines espèces apparaissent
extrêmement vulnérables, ce qui signifie qu’elles seraient exposées
quotidiennement à ces embolies, en milieu de journée par exemple. Ce type de
fonctionnement est très surprenant, c’est un peu se couper une jambe pour
courir plus vite ! Par ailleurs les conditions de recouvrements nocturnes
de ces plantes restent ‘miraculeuses’. Dans cette étude, nous avons étudié en
détail ces différentes techniques pour un grand nombre d’espèces. Nous sommes
arrivés à la conclusion que les techniques récentes sont fortement biaisées
pour certaines espèces, celles dont le tissu vasculaire est formé de grands
vaisseaux (comme les chênes ou la vigne par exemple). Nos travaux remettent
clairement en cause de nombreuses conclusions récentes tirées à partir de
résultats expérimentaux biaisés.
Dans les plantes,
la sève circule sous tension, c'est-à-dire sous des pressions négatives !
Cette état de l’eau est physiquement instable et peut évoluer vers une phase
gazeuse, plus stable. C’est le phénomène de cavitation. Il provoque une embolie
vasculaire qui bloque la circulation de la sève. C’est une perturbation majeure
car elle peut conduire à la mort des plantes (voir la brève de Janvier 2009
ci-dessous). De nombreuses techniques ont été proposées pour étudier la
cavitation et l’embolie des plantes et notre laboratoire a été particulièrement
pionnier dans ce domaine. Les techniques ‘anciennes’ sont fiables, mais peu
rapides. Elles ont néanmoins permis de faire considérablement progresser notre
compréhension du fonctionnement hydrique des plantes (dans le domaine c’est
probablement l’une des avancées majeures des 2 dernières décennies, avec la
découverte des aquaporines). Les
résultats acquis avec ces méthodes suggèrent que la cavitation (estivale) est
un phénomène exceptionnel qui se produit lorsque les plantes atteignent leur
limite de fonctionnement, par exemple lors d’une sécheresse d’une grande
ampleur (1976, 2003). On pense donc que c’est un processus impliqué dans
la survie ou la résilience des espèces à ces sécheresses. Les techniques
‘modernes’ (telle la centrifugation ou l’injection d’air) sont beaucoup plus
rapides. Certaines, comme le ‘Cavitron’ que nous avons développé au
laboratoire, permettent même de faire du phénotypage
à moyen débit de la cavitation. Ces techniques sont maintenant utilisées en
routine dans de nombreux laboratoires dans le monde. Pour certaines espèces,
les résultats obtenus avec ces techniques changent radicalement notre
perception du phénomène de cavitation. En effet, certaines espèces apparaissent
extrêmement vulnérables, ce qui signifie qu’elles seraient exposées
quotidiennement à ces embolies, en milieu de journée par exemple. Ce type de
fonctionnement est très surprenant, c’est un peu se couper une jambe pour
courir plus vite ! Par ailleurs les conditions de recouvrements nocturnes
de ces plantes restent ‘miraculeuses’. Dans cette étude, nous avons étudié en
détail ces différentes techniques pour un grand nombre d’espèces. Nous sommes
arrivés à la conclusion que les techniques récentes sont fortement biaisées
pour certaines espèces, celles dont le tissu vasculaire est formé de grands
vaisseaux (comme les chênes ou la vigne par exemple). Nos travaux remettent
clairement en cause de nombreuses conclusions récentes tirées à partir de
résultats expérimentaux biaisés.
Illustration : Image en microscopie électronique à balayage de deux
ponctuations dans le bois de cèdre. Les ponctuations (pits
en anglais) sont le lieu de passage de la sève brute d’un conduit à l’autre du
tissu vasculaire des plantes. C’est aussi le lieu supposé d’entrée d’air lors
du phénomène de cavitation. Les méthodes récentes de mesure de la cavitation
réservent des pièges (pitfall en anglais) dans
lesquels il ne faut pas tomber ! (photo : H Cochard)
Publication: Cochard H, Herbette S, Barigah T, Badel E, Ennajeh M, Vilagrosa A. (2010). Does sample length influence the shape of xylem embolism
vulnerability curves? A test with the Cavitron spinning
technique. Plant Cell and Environment 33: 1543–1552. pdf
Mars
2010 : La nervation des feuilles dévoile ses secrets.
Les feuilles des plantes à fleurs (Angiospermes) sont
parcourues par un réseau extrêmement dense de nervures qui irriguent toute leur
surface. Ce système d’irrigation est bien plus performant que celui des autres
espèces. Cela leur permet alors de développer un très grand nombre de pores à
la surface de leurs feuilles (les stomates), pores par lesquels entre le gaz
carbonique nécessaire à la photosynthèse mais aussi par où s’échappe
inéluctablement de grandes quantités d’eau. La nervation des feuilles est
extraordinairement diverse dans le monde végétal. Pour comprendre cette
diversité, nous avons construit un modèle informatique sophistiqué représentant
aussi fidèlement que possible les centaines de milliers de nervures qui
parcourent une feuille. Le modèle calcul les flux de sève et les pertes de
charges dans chacune des ramifications de ce système d’irrigation ce qui permet
d’apprécier sa performance hydraulique globale. Le modèle calcul également de
cout de construction en énergie de la nervation, les processus évolutifs devant
favoriser les solutions les plus efficaces au moindre cout. Nous avons alors
manipulé les paramètres du modèle de façon à identifier ceux qui confèrent au
réseau la meilleure performance hydraulique au meilleur cout énergétique. Le
fait que les nervures soient toutes interconnectées augmente considérablement
la performance hydraulique. La densité des nervures et la performance
hydraulique de chaque nervure sont également des éléments contribuant très
favorablement à la performance du système. Dès lors, on pourrait penser
que la sélection favoriserait les feuilles ayant une très forte densité de
grosses nervures très performantes. Il n’en est rien. La raison principale est
que une telle nervation aurait un cout de construction beaucoup trop élevé au
regard du gain fonctionnel acquis. Les angiospermes ont trouvé deux parades
pour diminuer ce cout de construction. Tout d’abord, la nervation est
hiérarchisée : un petit nombre de grosses nervures principales irriguent
un plus grand nombre de veines secondaires plus fines qui elle-même alimentent
une innombrable quantité de veinules très fines. Ensuite, les nervures principales
et secondaires ont une taille qui diminue progressivement de la base vers leur
extrémité. Ce défilement traduit le fait que la base de la nervure irrigue une
plus grande surface de la feuille que son extrémité. Ces deux propriétés de la nervation des feuilles
diminuent considérablement leur cout de construction tout en maintenant des
performances hydrauliques élevées.
A la suite de notre publication, Beerling
et Franks (Nature, 2010) ont trouvé dans nos travaux
une explication rationnelle au succès de l’évolution des Angiospermes depuis le
Crétacé. Avant le Crétacé, la planète était dominée par des plantes comme les
Gymnospermes dont le système hydraulique foliaire était peu performant.
Néanmoins, à cette époque la concentration atmosphérique en gaz carbonique
était bien supérieure à la concentration actuelle, ce qui signifie qu’une
faible densité de stomates suffisait à alimenter en carbone les feuilles. A la
fin du Crétacé (il y a 130 millions d’années) la concentration en CO2 chute
progressivement ce qui favorisa les plantes ayant des densités stomatiques
fortes et donc des taux de photosynthèse élevés. Inévitablement, ceci c’est
aussi traduit par une augmentation progressive des pertes en eau des feuilles
et donc par une pression de sélection forte pour les espèces ayant des systèmes
vasculaires plus performants. Seuls les Angiospermes avec leur système
vasculaire réticulé ont su rester dans cette course évolutive. Elles ont
progressivement augmenté la densité de leur fines nervures, accentué le défilement
des nervures principales et développé des nervations hiérarchisés. Comme le
démontre notre étude, ces innovations ont permis une meilleure performance
hydraulique, une minimisation du cout énergétique de construction pour une
amélioration des capacités photosynthétiques foliaires.
Illustration : Nervures d’une feuille de Noyer. Ces nervures
constituent un système d’irrigation en eau des feuilles dont les
caractéristiques maximisent la performance hydraulique tout en minimisant le
cout de construction. Une prouesse remarquable qui a certainement contribuée au
succès des plantes à fleurs depuis la fin du Crétacé. (photo H. Cochard)
Publication: McKown AD, Cochard H, Sack L (2010) Decoding leaf hydraulics with a spatially
explicit model: Principles of venation architecture and implications for its
evolution. The American Naturalist 175 : 447–460
Voir aussi en fin d’article le News
& Views de Beerling DJ and Franks (2010). The hidden cost of transpiration. Nature 464: 495-496.
Octobre
2009 : Un nouveau mécanisme explique la cavitation chez les conifères.
 Le risque de cavitation des
cellules conductrices de sève brute est un risque majeur pour la survie des
plantes car les cellules cavités deviennent impropres à la conduction de l’eau.
Beaucoup d’espoirs sont mis actuellement dans l’utilisation de ce trait de
fonctionnement comme marqueur de la résistance à la sécheresse des
arbres. Néanmoins ces espoirs sont pour le moment freinés par notre manque
de connaissance sur les mécanismes de formation de la cavitation et sur les
gènes qui les contrôlent. L’objectif de ce travail était d’élucider ces
mécanismes chez les conifères. Nous savons que la cavitation se forme suite à
l’entrée d’une bulle d’air dans le tissu conducteur. C’est le mécanisme du
« germe d’air ». Les sites les plus probables de cette entrée d’air
sont les ponctuations. Ce sont de minuscules pores dans la paroi des conduits
qui permettent le passage de la sève brute d’un conduit à l’autre. Chez les
conifères, comme les pins, les sapins ou les cèdres, ces ponctuations ont une
forme spectaculaire de trampoline ou de gong (cf la
photo de la brève d’Avril 2010 ci-dessus). La partie centrale de la membrane
interne de la ponctuation (le torus) est pleine alors que sa partie
périphérique est formée de fine fibrilles (la marge). Cette membrane fonctionne
comme une minuscule soupape. Lorsque les conduits de part et d’autre de la
ponctuation sont pleins de sève, la membrane se situe au milieu de la
ponctuation et la sève circule à travers les fibres de la marge. Par contre,
lorsqu’un des conduit cavite, le torus se déplace
vers la paroi et opercule son ouverture. L’air ne peut alors pénétrer dans le
second conduit, confinant ainsi l’embolie. On pensait jusqu’à maintenant que la
propagation de la cavitation se faisait par rupture des microfibrilles
de la marge (cas B) ou par glissement du torus (cas C). Sous ces hypothèses, ce
sont les propriétés mécaniques des microfibrilles qui
seraient en jeu dans le mécanisme. Nous avons émis une hypothèse alternative
(cas D) : la cavitation serait liée à un défaut d’étanchéité de la soupape
et serait donc causé par le passage d’une bulle d’air au niveau du joint entre
le torus et la paroi, voir même à travers le torus. Du point de vue physique,
on serait en présence d’une rupture capillaire d’un ménisque. Pour tester cette
hypothèse, nous avons fait varier la tension de surface de la sève (par
adjonction de solvant) de façon à élever le point de rupture capillaire. Nos
résultats confirment cette hypothèse : plus la tension de surface diminue,
plus la cavitation est facilitée. Le mécanisme de cavitation chez les conifères
est donc très plus proche de celui des autres espèces (comme les Angiospermes)
qui ont des structures anatomiques très différentes (pas de torus par exemple).
En d’autres termes, il est possible que les bases génétiques qui gouvernent la
cavitation soient communes pour toutes ces espèces. Mais ce point reste à
démontrer.
Le risque de cavitation des
cellules conductrices de sève brute est un risque majeur pour la survie des
plantes car les cellules cavités deviennent impropres à la conduction de l’eau.
Beaucoup d’espoirs sont mis actuellement dans l’utilisation de ce trait de
fonctionnement comme marqueur de la résistance à la sécheresse des
arbres. Néanmoins ces espoirs sont pour le moment freinés par notre manque
de connaissance sur les mécanismes de formation de la cavitation et sur les
gènes qui les contrôlent. L’objectif de ce travail était d’élucider ces
mécanismes chez les conifères. Nous savons que la cavitation se forme suite à
l’entrée d’une bulle d’air dans le tissu conducteur. C’est le mécanisme du
« germe d’air ». Les sites les plus probables de cette entrée d’air
sont les ponctuations. Ce sont de minuscules pores dans la paroi des conduits
qui permettent le passage de la sève brute d’un conduit à l’autre. Chez les
conifères, comme les pins, les sapins ou les cèdres, ces ponctuations ont une
forme spectaculaire de trampoline ou de gong (cf la
photo de la brève d’Avril 2010 ci-dessus). La partie centrale de la membrane
interne de la ponctuation (le torus) est pleine alors que sa partie
périphérique est formée de fine fibrilles (la marge). Cette membrane fonctionne
comme une minuscule soupape. Lorsque les conduits de part et d’autre de la
ponctuation sont pleins de sève, la membrane se situe au milieu de la
ponctuation et la sève circule à travers les fibres de la marge. Par contre,
lorsqu’un des conduit cavite, le torus se déplace
vers la paroi et opercule son ouverture. L’air ne peut alors pénétrer dans le
second conduit, confinant ainsi l’embolie. On pensait jusqu’à maintenant que la
propagation de la cavitation se faisait par rupture des microfibrilles
de la marge (cas B) ou par glissement du torus (cas C). Sous ces hypothèses, ce
sont les propriétés mécaniques des microfibrilles qui
seraient en jeu dans le mécanisme. Nous avons émis une hypothèse alternative
(cas D) : la cavitation serait liée à un défaut d’étanchéité de la soupape
et serait donc causé par le passage d’une bulle d’air au niveau du joint entre
le torus et la paroi, voir même à travers le torus. Du point de vue physique,
on serait en présence d’une rupture capillaire d’un ménisque. Pour tester cette
hypothèse, nous avons fait varier la tension de surface de la sève (par
adjonction de solvant) de façon à élever le point de rupture capillaire. Nos
résultats confirment cette hypothèse : plus la tension de surface diminue,
plus la cavitation est facilitée. Le mécanisme de cavitation chez les conifères
est donc très plus proche de celui des autres espèces (comme les Angiospermes)
qui ont des structures anatomiques très différentes (pas de torus par exemple).
En d’autres termes, il est possible que les bases génétiques qui gouvernent la
cavitation soient communes pour toutes ces espèces. Mais ce point reste à
démontrer.
Illustration : Coupes transversale d’une ponctuation de conifère
illustrant les différents mécanismes possibles pour la cavitation. La
cavitation peut se produire soit par le passage d’une bulle d’air à travers les
microfibrilles de la marge (A), soit par la rupture
de ces fibrilles (B) ou le glissement du torus (C). Nous avons proposé une
hypothèse alternative : le passage d’une bulle d’air entre le torus et la
paroi, voire directement à travers le torus (D). Nos expérimentations sont
cohérentes avec ce nouveau mécanisme. (Dessin : H Cochard)
Publication: Cochard H, Holtta, Herbette S, Delzon S, Mencuccini M. 2009 New Insights into the Mechanisms of
Water-Stress Induced Cavitation in Conifers. Plant Physiology 2:
949-954
Janvier
2009 : On comprend maintenant mieux pourquoi les arbres plus résistants à
la cavitation sont aussi plus résistants à la sécheresse
 La durabilité des écosystèmes
forestiers est fortement compromise par l'occurrence d'événements climatiques
extrêmes tels que des sécheresses d'intensités exceptionnelles par exemple. La
fréquence de ces accidents risquant d'augmenter dans le futur, il importe de
comprendre les mécanismes de survie des espèces forestières afin d'identifier
du matériel génétique plus performant. La résistance du système vasculaire à la
cavitation est un paramètre fortement corrélé aux exigences en eau des essences
forestières et pourrait donc constituer un critère de sélection efficace pour
la résistance à la sécheresse des arbres. Le but de ce travail était donc
d'identifier des mécanismes physiologiques permettant d’expliquer la relation
entre résistance à la cavitation et résistance à la sécheresse.
La durabilité des écosystèmes
forestiers est fortement compromise par l'occurrence d'événements climatiques
extrêmes tels que des sécheresses d'intensités exceptionnelles par exemple. La
fréquence de ces accidents risquant d'augmenter dans le futur, il importe de
comprendre les mécanismes de survie des espèces forestières afin d'identifier
du matériel génétique plus performant. La résistance du système vasculaire à la
cavitation est un paramètre fortement corrélé aux exigences en eau des essences
forestières et pourrait donc constituer un critère de sélection efficace pour
la résistance à la sécheresse des arbres. Le but de ce travail était donc
d'identifier des mécanismes physiologiques permettant d’expliquer la relation
entre résistance à la cavitation et résistance à la sécheresse.
Pour cette étude nous avons sélectionné quatre espèces de
conifères de l’hémisphère sud ayant des préférences écologiques très
contrastées (des forêts humides de Tasmanie aux zones semi-arides du sud-est de
l'Australie), la résistance à la cavitation des espèces des zones sèches étant
considérablement plus grandes. Les plantes ont été élevées en pots, exposées à
des sécheresses d'intensités croissantes, puis réhydratées. Comme attendu, les
espèces les plus résistantes à la cavitation ont survécu à des intensités de sécheresse
bien plus élevées. Pour la première fois, nous avons démontré que l'induction
de cavitation dans les tiges et les aiguilles de ces espèces altérait leur
capacité de récupération après réhydratation. Un arbre dont le système
vasculaire reste intègre suite à une sécheresse retrouve un fonctionnement
physiologique normal après une nuit. En revanche, pour les individus ayant
perdu une partie de la fonctionnalité de leurs vaisseaux, cette période
augmente rapidement pour devenir infinie pour des taux de cavitation proche 50%
dans les tiges.et de 95% dans les aiguilles Posséder un système vasculaire plus
résistant à la cavitation est donc un caractère adaptatif pour les espèces
colonisant les milieux arides.
Les résultats de cette expérimentation conduite sur des
espèces de conifères sont en cours de validation pour des espèces de feuillus
dans notre laboratoire. En parallèle, nous avons entrepris la caractérisation
de la résistance à la cavitation de l'ensemble des essences forestières de la
flore française. Par ailleurs nous cherchons maintenant à identifier les bases
génétiques de la cavitation sur des espèces modèles. Ceci nous permettra, à
terme, de fournir des critères objectifs de sélection pour l'amélioration de la
résistance à la sécheresse de nos espèces forestières. Ce travail est le fruit
d'une collaboration avec Tim Brodribb de l'Université
de Tasmanie en Australie.
Illustration : Un Hêtre mort, peut-être à cause d’une sécheresse? Les
mécanismes qui conduisent à la mort d’un arbre à la suite d’une sécheresse
restent méconnues. Nous montrons dans cette étude que les phénomènes de
cavitation sont l’une des clés de cette compréhension. (photo : H. Cochard)
Publication: Brodribb T. and Cochard H. 2009. Hydraulic Failure Defines the Recovery and Point of
Death in Water-Stressed Conifers. Plant Physiology 149 : 575-584
Janvier
2007 : Des protéines (aquaporines) contrôlent les flux d’eau dans les
feuilles des arbres.
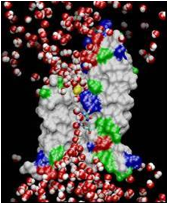 Les échanges d’eau et de carbone entre les arbres et l’atmosphère
s’effectuent principalement au niveau des feuilles. Déterminer les bases
physiologiques, biophysiques et moléculaires de ces échanges gazeux est
nécessaire pour comprendre l’adaptation des arbres aux fluctuations de leur
environnement.
Les échanges d’eau et de carbone entre les arbres et l’atmosphère
s’effectuent principalement au niveau des feuilles. Déterminer les bases
physiologiques, biophysiques et moléculaires de ces échanges gazeux est
nécessaire pour comprendre l’adaptation des arbres aux fluctuations de leur
environnement.
Avant d’être rejetée dans l’air sous forme gazeuse, la sève brute
circule sous forme liquide dans l’arbre dans des structures anatomiques très
spécialisées. Il est établi que la facilité de circulation de la sève dans les
feuilles conditionne les quantités d’eau et carbone que les arbres échangent
avec l’atmosphère. Le réseau extrêmement dense de fines nervures qui irrigue toute
la feuille rend très efficace le transport de la sève sur la quasi-totalité de
son trajet foliaire. Le goulot d’étranglement de circulation de l’eau dans la
feuille se localise sur une distance très courte, entre la terminaison des
nervures et les sites d’évaporation. Il était jusqu’à présent admis que l’eau
circulait ici principalement dans la paroi inerte des cellules. Or nous avons
démontré que cette circulation répondait rapidement et fortement à des
variations de température, de luminosité et à la présence de certaines drogues.
Ceci démontre l’existence d’un contrôle biologique de la circulation de l’eau
dans la feuille impliquant un flux d’eau au travers des membranes plasmiques et
non plus au travers des parois cellulaires. Les bases moléculaires de ce trajet
ont été identifiées : il s’agit de protéines ‘canaux à eau’ trans-membranaires
(aquaporines) ayant la capacité de s’ouvrir ou de se fermer. Les arbres ont
donc la faculté d’ajuster la capacité de leurs feuilles à transporter l’eau en
fonction des conditions environnementales. Des modèles suggèrent que ce
processus permettrait d’optimiser leurs échanges gazeux et croissances.
L’identification des processus et des bases moléculaires qui contrôlent des
flux d’eau dans l’arbre fournira des outils innovants pour la sélection de
génotypes plus productifs ou plus économes en eau. Cette étude a été menée en
collaboration avec le Pfr Melvin Tyree
de l’Edmonton University au Canada.
Illustration :
Modèle numérique d’une aquaporine. Les aquaporines
sont des protéines qui laissent passer les molécules d’eau à travers la
membrane plasmique des cellules. Notre étude démontre leur rôle dans la
régulation des flux d’eau dans les feuilles. (photo :http://nobelprize.org/chemistry/laureates/2003/chemanim1.mpg)
Publication: Cochard H, Venisse JS, Barigah TS, Brunel N, Herbette S,
Guilliot A, Tyree MT, Sakr
S 2007 Putative Role of
Aquaporins in Variable Hydraulic Conductance of Leaves in Response to
Light. Plant Physiology 143:122-13